Fluorescent In-Situ Hybridization (FISH)
Fluorescent In-Situ Hybridization is a process which vividly paints chromosomes or portions of chromosomes with fluorescent molecules.
First introduced in the early 1980s and considered to be a new advent in the field of cytology, FISH (Figure 1) is a technology useful for identifying chromosomal abnormalities and for gene mapping.

Figure 1. Fluorescence In-Situ Hybridization (National human genome research institute, 1).
Fluorescence In-Situ Hybridization is commonly used to effectively and efficiently detect oncogenes in tumours, including both haematological malignancies and solid tumours, and diagnose congenital (such as Down's and Edward's Syndrome) or infectious diseases, increasing its worldwide application trends in recent years.
Compared to a normal application to clinical diagnosis, the level of accuracy and specificity of these diagnoses by FISH is higher and more reliable than other molecular profiling techniques, e.g. single nucleotide polymorphism (SNP) or array-based comparative genomic hybridization, and therefore plays a key role in the prevention, treatment, and reduction of pain and agony in patients, as well as paving the way for further clinical developments in these fields of medical science. As an example, the resolution of FISH is 50 times greater than that of Giemsa staining conventionally used to detect genetic diseases. Besides, it is considerably less tedious in terms of time than karyotyping, which necessitates cell culture and successive metaphase arrest to examine the structural characteristics of chromosomes.
Albeit FISH has very limited use in developing countries due to the non-availability and insufficient expertise, it has received great recognition as a physical mapping technique to support large-scale mapping and sequencing endeavours related to the human genome project.
The basis of FISH is the high affinity with which complementary base pairs (A-T, C-G) bind to their corresponding partner, which is called „hybridization“. FISH is performed with either a directly or indirectly labelled hybridizing DNA probe. Direct labelling uses fluorescent nucleotides, while indirect labelling is embedded with reporters that are successively detected by fluorescent antibodies or other affinity molecules.
This extraordinary technique offers an intermediate degree of resolution between DNA analysis and chromosomal investigations. Fluorescent DNA probes, attached to complementary parts of the chromosome, emit coloured signals that can be easily picked up and visualized. Should there have been a duplication, more of the probe will hybridize with the sample DNA and greater fluorescence will be revealed. A deletion in the genome prevents the probe from attaching to its missing counterpart region and thus fluorescence will not be detected. Consequently, diverse types of FISH probes can detect different chromosome aberrations depending on the specificity of the probes for a particular gene locus.
The protocol normally consists in the preparation of the fluorescent probes, followed by denaturation of both target and probe DNA strands and their consequent re-hybridization for subsequent detection (Figure 2).
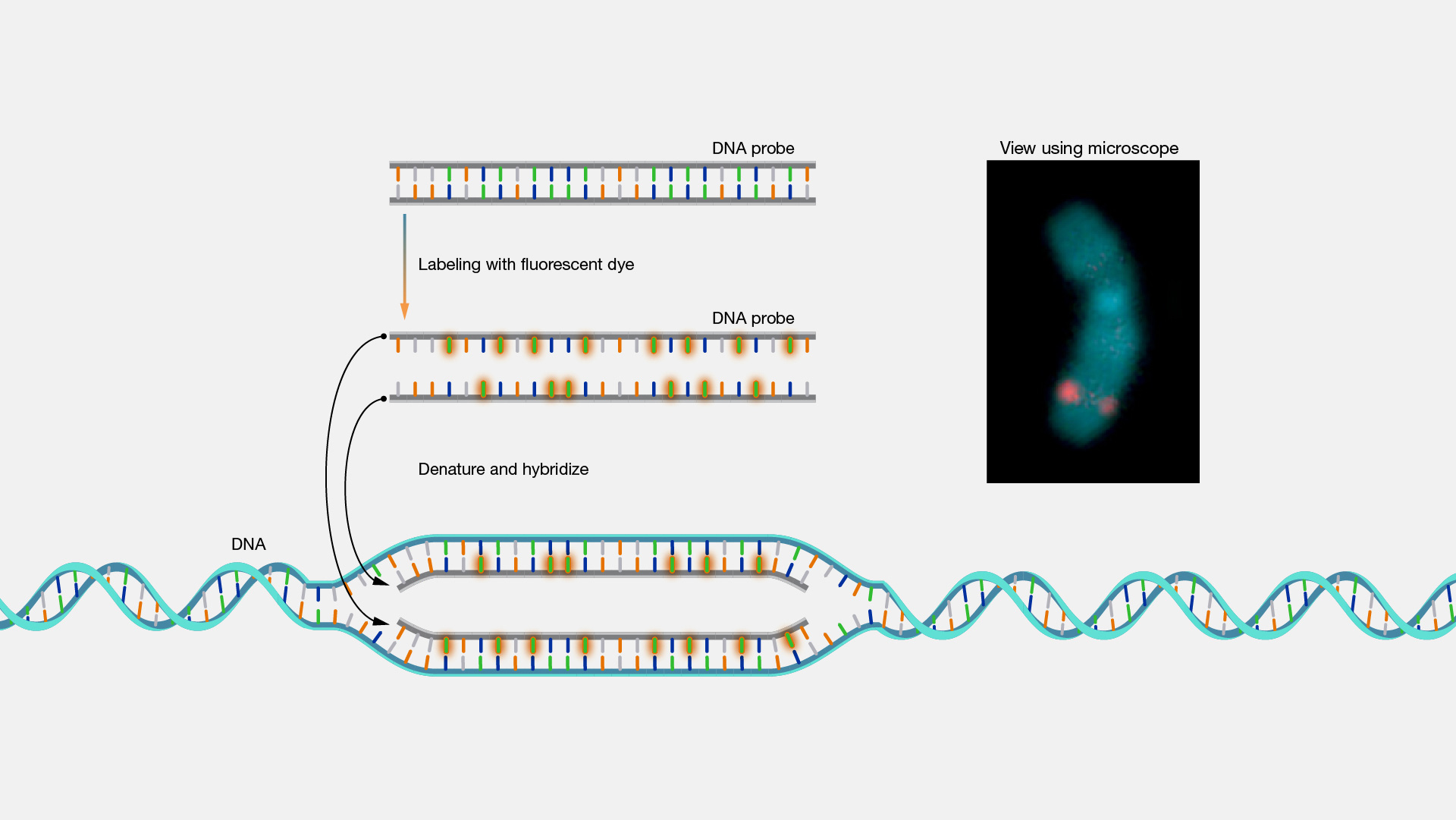
Figure 2. Basic principle of Fluorescence In-Situ Hybridization (National human genome research institute, 2).
As shorter probes hybridize less specifically than longer ones, probe size is important, so rather long DNA or RNA strands (often 10-25 nucleotides) that are complementary to a given target sequence are often used to locate a target. Complementarity sets the resolution of the characteristics that can be detected. For example, for an experiment to determine the breakpoint of a translocation, probes' overlap specifies the narrow window in which the breakpoint can be found.
In "whole chromosome painting", whole chromosome hybridization probes are used to count the number of a given chromosome, display translocations or pinpoint extrachromosomal chromatin fragments.
A multitude of techniques use a series of probes of different colours.
The comparative genomic hybridization (CGH) technique involves two-colour FISH, which is useful for establishing the ratios of fluorescence intensity values between tumour and control DNA along normal reference chromosomes in metaphase, and thus for identifying DNA copy number alterations. Probe mix ratios can be used to create secondary colours. Similar to comparative genomic hybridization, probe mixes are created in M-FISH by mixing the correct ratio of two sets of probes of different colours for the same chromosome. ). More recently, each homologous chromosome pair in M-FISH is uniquely labelled with five sets of fluorochromes with different combinations that are spectrally distinct. Certain tests are arranged so that there is either a secondary colour present or absent in the cases of interest. For example, for translocation detection, when immediately contiguous colours appear to overlap, a secondary colour is observed. In double-fusion FISH or D-FISH, the secondary colour detects the disease (e.g. the detection of BCR/ABL1 translocations in chronic myeloid leukemia), while in the so-called "break-apart FISH", the lack of the secondary color is pathological. Indeed, secondary colours are observed in normal cells, whereas when translocation occurs, only primary colours are observed.

Figure 3. SKY image showing metaphase chromosomes labeled with different fluorochromes (3).
A new molecular cytogenetic method to characterise numerical and structural chromosome aberrations is spectral karyotyping (SKY, Figure 3), which involves the concurrent hybridization of 24 differentially labelled chromosome probes and generates a full-genome colour karyotype.
Together with multicolour FISH, SKY enables all chromosomes to be displayed at the same time by „painting“ them with a combination of different colours that are spectrally distinguishable fluorochromes, but different methods are used to reveal and differentiate the various fluorescence combinations after in-situ hybridization.
The use of SKY has contributed significantly to the improvement oft he identification of chromosomal abnormalities associated with constitutional cytogenetics and oncology.
New and exciting applications of FISH are being developed that broaden its scope. Complementary 16S rRNA oligonucleotide sequences (17-34 nucleotides long) as FISH probes can be employed for the investigation of microbial communities both within the oral cavity and the gastrointestinal flora, and even detect malaria infections in blood samples. Targeted oligonucleotide probes have also been engineered to identify disease agents in respiratory tract infections. While recently, researchers have succeeded in boosting the resolution of FISH by targeting elongated chromatin fibres or microarrays.
At metabion, we are proud to provide our customers with top quality products, developed and produced using state-of-art technology. An excellent example is our synthesis service for custom FISH probes, which offers significantly increased flexibility in terms of
- multiple labeling of custom FISH probes (2 or more modifications)
- production of modified probes exceeding the critical sequence length imposed by conventional oligonucleotide synthesis (longmers).
Visit our product portfolio for Longmers and do not hesitate to ask for non-listed combinations including internally or multi-labeled oligos and probes.
For more information and a quotation, contact us at info@metabion.com!
References:
- National human genome research institute (https://www.genome.gov/genetics-glossary/Fluorescence-In-Situ-Hybridization).
- National human genome research institute (https://www.genome.gov/sites/default/files/media/images/tg/FISH.jpg).
- Chowdhury MR., Dubey S. Chapter 24 - Role of Cytogenetics and Molecular Genetics in Human Health and Medicine. Models in Discovery and Translation. Animal Biotechnology. 2020;451–472.
- Speicher MR., Carter NP. The new cytogenetics: blurring the boundaries with molecular biology. Nat Rev Genet. 2005 Oct;6(10):782-92.
- Schröck E., du Manoir S., Veldman T., Schoell B., Wienberg J., Ferguson-Smith MA., Ning Y., Ledbetter DH., Bar-Am I., Soenksen D., Garini Y., Ried T. Multicolor spectral karyotyping of human chromosomes. Science. 1996 Jul 26;273(5274):494-7.
- Speicher MR., Gwyn Ballard S., Ward DC. Karyotyping human chromosomes by combinatorial multi-fluor FISH. Nat Genet. 1996 Apr;12(4):368-75.
- Tkachuk DC., Westbrook CA., Andreeff M., Donlon TA., Cleary ML., Suryanarayan K., Homge M., Redner A., Gray J., Pinkel D. Detection of bcr-abl fusion in chronic myelogeneous leukemia by in situ hybridization. Science. 1990 Oct 26;250(4980):559-62.
- Parra I., Windle B. High resolution visual mapping of stretched DNA by fluorescent hybridization. Nat Genet. 1993 Sep;5(1):17-21.
- Ratan ZA., Zaman SB., Mehta V., Haidere MF., Runa NJ., Akter N. Application of Fluorescence In Situ Hybridization (FISH) Technique for the Detection of Genetic Aberration in Medical Science. Cureus. 2017 Jun 9;9(6):e1325.
- Lander ES. et al. International Human Genome Sequencing Consortium. Initial sequencing and analysis of the human genome. Nature. 2001 Feb 15;409(6822):860-921.
- Mei R., Galipeau PC., Prass C., Berno A., Ghandour G., Patil N., Wolff RK., Chee MS., Reid BJ., Lockhart DJ. Genome-wide detection of allelic imbalance using human SNPs and high-density DNA arrays. Genome Res. 2000 Aug;10(8):1126-37.
- McNeil N., Ried T. Novel molecular cytogenetic techniques for identifying complex chromosomal rearrangements: technology and applications in molecular medicine. Expert Rev Mol Med. 2000 Sep 14;2000:1-14.
- Jalal, S.M., Law, M.E. Multicolor FISH. In: Fan, YS. Molecular Cytogenetics. Methods in Molecular Biology. 2002;204:105-20. Humana Press.
- Peterson S., Rehen S., Westra W., Yung Y., Chun J. Spectral karyotyping and fluorescent in situ hybridization. In: Loring JF., Wesselschmidt RL., Schwartz PH. Human Stem Cell Manual: A Laboratory Guide. 2007;71–84.